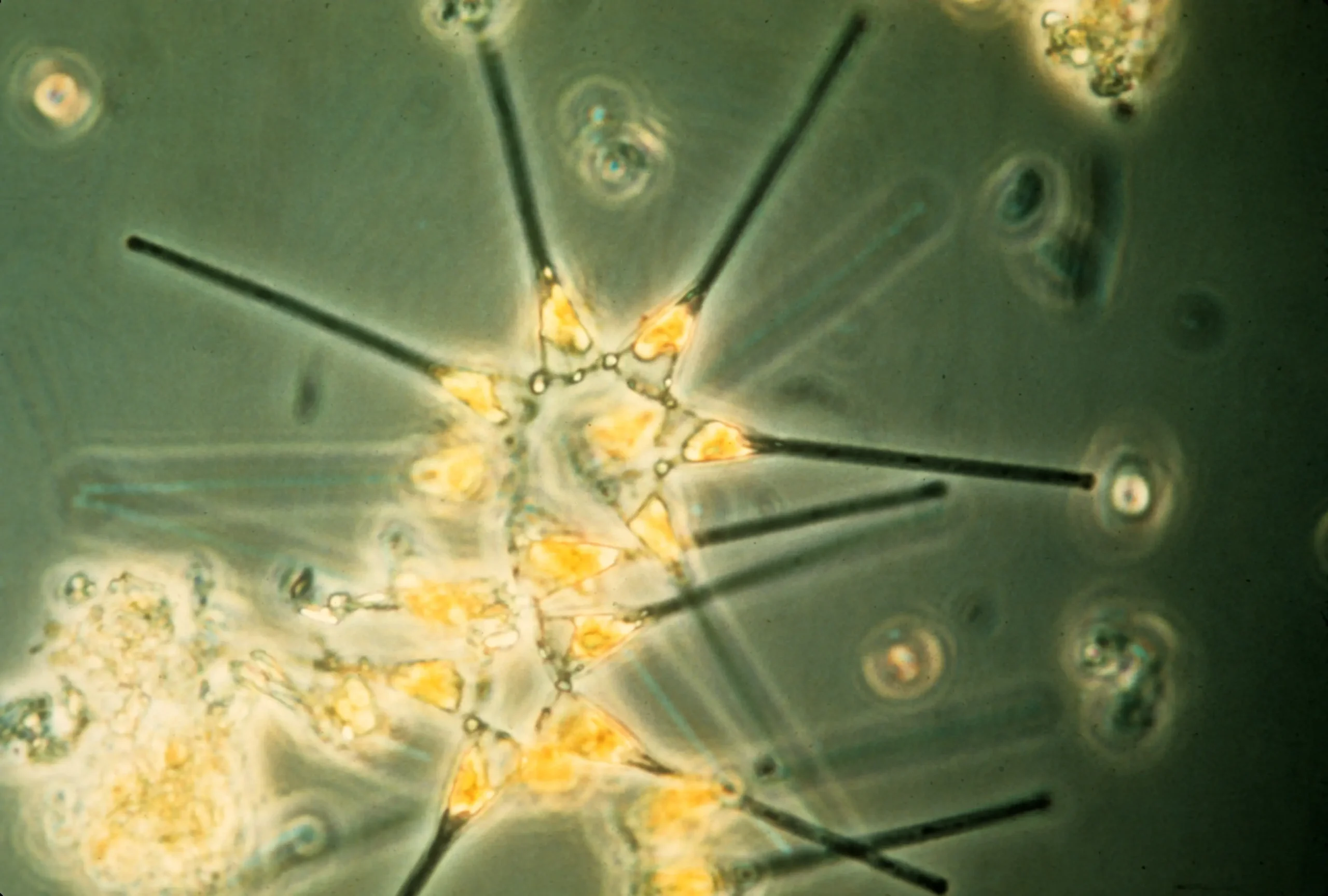
Marine Phytoplankton
Marine phytoplankton are microscopic, photosynthetic organisms that form the foundation of the ocean's food web and produce over half of Earth's oxygen.
Like land plants, phytoplankton use sunlight to convert carbon dioxide (CO2) and water into organic carbon for their body parts and release oxygen as a byproduct of photosynthesis. CO2 is therefore an essential nutrient for phytoplankton and higher concentrations of CO2 in the surface ocean have the potential to support higher rates of phytoplankton photosynthesis and growth.
Phytoplankton are essential for removing CO2 from the water and supplying O2, however, this balance can be disturbed when there is excess CO2 in the water.

The impacts of acidification to phytoplankton are highly complex and not well understood. With over 100,000 different species of phytoplankton, the responses of each individual species to acidification are highly varied and challenging to pinpoint. In laboratory experiments with pure cultures of individual species of phytoplankton, little or no change in growth is typically observed in cultures grown with CO2 levels expected in the year 2100.
However, other more subtle effects have been noted including interactions of elevated CO2 with demand for other nutrients such as nitrogen, phosphorus, and iron, which suggest that higher CO2 may be most beneficial to phytoplankton under nutrient rich conditions. Conversely, elevated CO2 can lead to reduced phytoplankton growth when occurring in combination with light or temperature stress.

While the effects of acidification on many phytoplankton species remain unclear, some information has been discovered about several key species important in coastal waters and central ocean basins.
Coccolithophores: In coccolithophores, a species of phytoplankton that produce calcium carbonate plates on their cell surface, decreases in pH can alter growth, calcification, and organic carbon production.
Trichodesmium: Lower pH may negatively affect the ability of Trichodesmium, an important species of photosynthetic cyanobacteria, to provide inorganic nitrogen to the ocean through nitrogen fixation, particularly in the vast areas of the ocean that are generally limited by nitrogen.

Harmful algal blooms (HABs) occur when there is rapid growth of certain species of phytoplankton, including Karenia brevis (red tide) and Aureococcus anophagefferens (brown tide), resulting in toxic conditions for both marine species and humans. Phytoplankton, such as Karenia brevis (red tide), tend to thrive in high CO2 conditions. Aureococcus anophagefferens is another HAB species that thrives under high nitrogen and phosphorus conditions, which often accompany high CO2 conditions. HABs are considered co-stressors to acidic conditions and can have compounding negative impacts on coastal habitat and animals. Some species of Karenia have been found as far north as New Jersey but are more common in southeastern US waters. As ocean water temperatures increase, it is expected that more of these species will be found farther north.

As multiple factors influence photosynthesis and growth of phytoplankton in the ocean, it is difficult to predict the effects of higher CO2 on the productivity of natural phytoplankton communities. For example, concurrent increases in both CO2 and temperature could enhance phytoplankton or macroalgae growth, however, as in the case of HABs, this increase in growth is not always good for the environment or animals. Similarly, increases in CO2 expected over the next 100 years may favor the growth of larger diatom species whose growth can be limited by lower CO2 and nutrient levels. Diatoms are known to require more nutrients and CO2 than other species. Increases in CO2 levels in the open ocean and coastal waters could lead to changes in phytoplankton community composition with potential secondary effects on biogeochemical cycles and food webs.